Attention and visual awareness
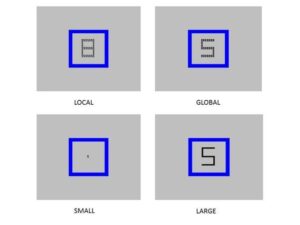
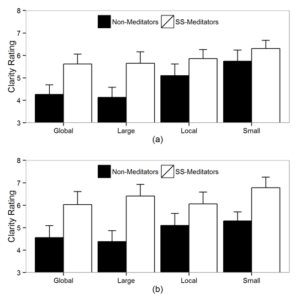
The research program on attention at CBCS includes studies on selective attention, attentional control, scope of attention, and attention-emotion interaction. There have been some projects investigating the effects of meditation on attention-mediated visual awareness. Using a paradigm involving color afterimages, the study showed that the practice of concentrative meditation results in larger afterimage duration and more clear afterimages (Srinivasan & Singh, 2017) (see Fig 1). The functional MRI study investigating the areas involved in the formation of afterimages showed larger activation in the right inferior frontal cortex and right occipital cortex (see Fig 2).
(a) (b)
Figure 1: (a) Stimuli used in afterimage study with meditators and non-meditators and (b) Clarity rating for SS-meditators and non-meditators under different load conditions.
[Reference: Srinivasan, N., & Singh, A. (2017). Concentrative meditation influences visual awareness: A study with color afterimages. Mindfulness, 8, 17-26]. 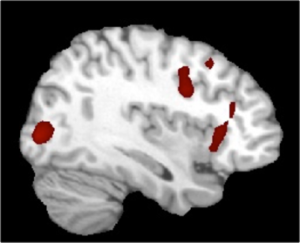
Figure 2: Activations for SS-meditators > non-meditators during afterimages
[Reference: Singh, A., Chandrasekhar, P. V., Guleria, A., Srinivasan, N. (2019). Concentrative (Sahaj Samadhi) meditation training and visual awareness: An fMRI study on color afterimages. Progress in brain research, 244, 185-206].
We have shown the effect of focused attention/concentrative meditation on time perception. We have also investigated the effect of scope of attention on time perception. With non-meditators, we see differences in duration estimation with larger slope for global compared to local processing. However, with meditators, local processing has a similar slope as global processing.
Self, Agency and Intentions
Intention and Perception
In a series of studies, we have investigated the effects of intentions on time and space perception (see Fig 3). We have shown even there is no predictability, forming an intention or desire to see a stimulus results in perceiving that stimulus faster and experiencing that stimulus as lasting longer compared to a stimulus one did not intend or desire to see (Makwana & Srinivasan, 2017).
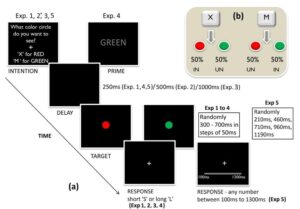
Figure 3: Trial structure for experiments on the effects of intention on time perception
[Reference: Makwana, N., & Srinivasan, N. (2017). Intended outcome expands in time. Scientific Reports, 7:6305].Emotion and intention
Another study at the Centre examined the role of emotion and action as a function of sense of agency informing the predictive models using the intentional binding paradigm. Results suggest a significant effect of intentional binding in favour of negative emotions and that sense of agency is prospective in nature in the context of emotional stimuli particularly when the outcome is unpredictable (Sarma & Srinivasan, 2021).
[Reference: Sarma, D., & Srinivasan, N. (2021). Intended emotions influence intentional binding with emotional faces: Larger binding for intended negative emotions. Consciousness and Cognition, 92, 103136].Time perception
We have performed studies using different emotions to investigate the effect of emotions on time perception. For example, we have found that emotions influence temporal resolution as indicated by differences in just noticeable difference obtained using a temporal order judgment task. Similar effects have also been found with changes in scope of attention resulting in changes in temporal resolution.
Emotion and Attention
Many studies at the Centre in the past have focused on the interaction between attention and emotion. Studies have used many paradigms including visual search, load, dwell time, hierarchical stimulus processing, attention network task.
Scope of Attention and Emotions
In the past few years, we have published multiple studies arguing for a link between broad scope of attention/global processing and happy emotions as well as narrow scope of attention/local processing and sad emotions. In addition, participants remember the sad faces presented during local processing compared to sad faces presented during global processing (Srinivasan & Gupta, 2011).

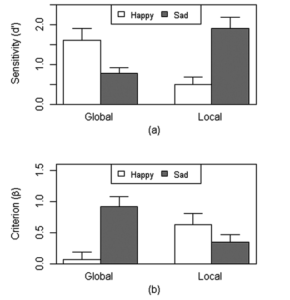
Figure 4: (a) A sample stimulus used to study effect of global-local processing on emotional processing and (b) sensitivity and criterion of recognition memory as a function of global-local processing and emotional content.
[Reference: Srinivasan, N., & Gupta, R. (2011). Global-local processing affects recognition of distractor emotional faces. Quarterly Journal of Experimental Psychology, 64, 425-433]Affective control: affective conflict adaptation
Our research program on affective control focuses on conflict monitoring, adaptation, and proactive control in task relevant positive and negative emotions using the face-word Stroop task (see Figure 5). Our studies suggest that affective conflict adaptation is modulated by the content of emotions with negative emotions recruiting greater proactive control driven by narrowing of attention and complexity of processing in case of negative emotion (see Figure 6) (Kar et al., 2018).
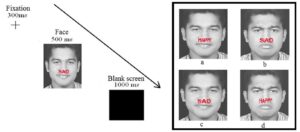
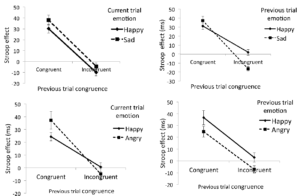
Figure 6. Affective adaptation as a function or previous trial congruence and emotion across positive and negative emotions.
[Reference: Kar, B. R., Srinivasan, N., Nehabala, Y., & Nigam, R. (2018). Proactive and reactive control depends on emotional valence: A Stroop study with emotional expressions and words. Cognition & Emotion, 32, 325-340.]Response inhibition in the presence of irrelevant positive and negative faces
This research explored the effect of positive and negative emotion perception on reactive inhibitory control. The emotional Stop-Signal task was administered to tap reactive inhibition. Emotional faces (positive, negative, and neutral) were presented as stop signals. This research indicates that approach and avoidance associated with emotional faces affect response inhibition.
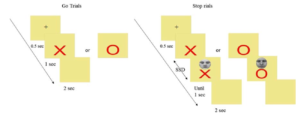
Figure 7: Trial structure for Emotional Stop-Signal task
[Reference: Gupta, R., Singh, J.P. (2021). Only irrelevant angry, but not happy, expressions facilitate the response inhibition. Attention, Perception, & Psychophysics, 83, 114–121. https://doi.org/10.3758/s13414-020-02186-w ]
In this line of research, we are further interested in examining the role of positive and negative emotion in proactive inhibitory control. In an ongoing study, we administered the emotional Go-NoGo task to examine the effect of positive and negative emotion on proactive inhibitory control.
Anxiety and cognitive control
One of our ongoing studies on trait anxiety and dynamic adjustments in control in healthy individuals using behavioural and EEG ERP methodology, as well as in those with generalized anxiety disorder (GAD) with the AX-CPT task demonstrate that trait anxiety affects processing efficiency and flexible cognitive control. Pathological anxiety (GAD) affects context processing required to exercise proactive control. In the past we have also investigated attentional control in the context of emotion stimuli in individuals with subclinical depression and anxiety.
Language and cognition
The overall goal of our research aims at exploring cognitive and neural mechanisms of language processing in its various manifestations.
Bilingualism and cognitive control
We study the interaction between bilingualism and cognitive control. We find that L1 proficiency extends itself to general-purpose cognitive control mechanisms whereas L2 proficiency contributes to bilingual language control. ERP studies using the visual identity negative priming paradigm with linguistic stimuli have shown early latencies and larger amplitudes for inhibitory control in case of L1 (see Figure 7) (Dash & Kar, 2020). We have also found default state proactive control and reduced proactive inhibition cost in high proficiency bilinguals using eye movement method (see Figure 8) (Singh & Kar, 2018). The sociolinguistic factors are found to moderate the interaction between bilingualism and inhibitory control (Thanissery, Parihar & Kar, 2020). We are currently investigating the neural mechanisms of the interaction between language and cognitive control networks using control tasks in fMRI.


Figure 8. Shows the sample stimuli (left) used in a visual identity negative priming paradigm to look at bilingual language control in bilinguals. Results showed greater amplitudes and earlier latencies of the N200 component for negative priming effect for L1 Hindi.
[Reference: Dash, T. & Kar, B. R. (2020). Behavioural and ERP correlates of bilingual language control and general-purpose inhibitory control predicted by L1 and L2 proficiency, Journal of Neurolinguistics, 56 doi.org/10.1016/j.jneuroling.2020. 100914].
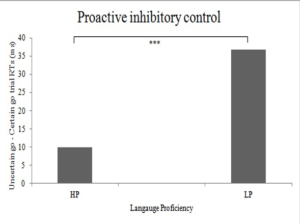
Figure 9. Reduced Proactive inhibitory cost in high-proficient bilinguals in a cued go/no-go task.
[Reference: Singh, J. P. Kar, B. R. (2018). Effect of language proficiency on proactive occulo-motor control among bilinguals. PloS one, 13(12), e0207904.Thanissery, N., Parihar, P., & Kar, B. R. (2020). Language Proficiency, sociolinguistic factors and inhibitory control. In the special issue on Cognitive Consequences of Bilingualism, Journal of Cultural Cognitive Science, 4 (2), DOI: 10.1007/s41809-020-00065-2]
A series of studies were conducted to examine if a bilingual’s ability in conflict resolution and inhibitory control as seen with manual tasks extends into other domains of human action control such as the oculomotor domain (Singh & Mishra, 2012).

Figure 10. Trial Sequence for Incongruent condition. Fixation proportions to the correct target and distractors for high and low proficient bilinguals for colour Stroop (panel A) and direction Stroop (panel B) trials.
[Reference: Singh, N., & Mishra, R. K. (2012). Does language proficiency modulate oculomotor control? Evidence from Hindi-English bilinguals. Bilingualism: Language and Cognition, 15, 771-781]Another study (Singh & Mishra, 2013) investigated the role of second language proficiency on the modulation of conflict-monitoring in bilinguals. The results support a conflict-monitoring account in a novel oculomotor task and also suggest that language proficiency could modulate executive control in bilinguals (Figure 10).
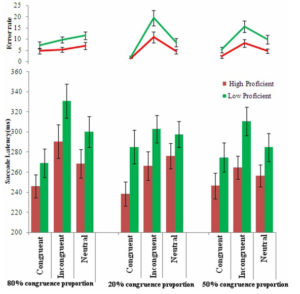
Figure 11: Sample trial (left). Mean saccade latency (ms) and error rates for high and low proficient bilinguals for the saccadic arrow Stroop task in different congruence proportion conditions (right).
[Reference: Singh, N., & Mishra, R. K. (2013). Second language proficiency modulates conflict monitoring in an oculomotor Stroop task: Evidence from Hindi-English Bilinguals. Frontiers in Psychology, 4:322]In a study by Singh & Mishra (2015), Hindi-English bilinguals differing in their L2 proficiency were compared on a saccadic countermanding task which taps inhibitory control as well as monitoring (see Fig 11) These results suggest that language proficiency may have an effect on performance monitoring, but not the inhibitory control per se (see Fig 12).
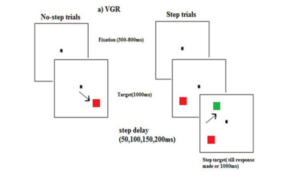
Figure 12: A sample trial sequence for step and no-step trials in the visually guided redirect (VGR) saccade task.
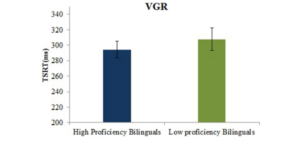
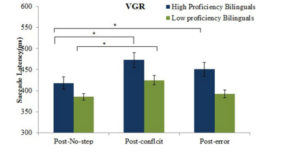
Figure 13: Mean TSRT (ms) for high and low proficient bilinguals on the VGR task (left). Mean saccade latencies to the post-conflict, post-error and post-no-step trials for high and low proficient bilinguals in the VGR task, ∗p < .05 (right).
[Reference: Singh, N., & Mishra, R. K. (2015). The modulatory role of second language proficiency on performance monitoring: evidence from a saccadic countermanding task in high and low proficient bilinguals. Frontiers in psychology, 5, 1481]Literacy and Cognition
This line of research explores the effect of reading and writing skills on the general cognitive process. In the past, Dr. Jay Prakash Singh was associated with the line of research work examining the interaction between literacy and cognition.
The effect of literacy on visual search
In this research, low and highly literate participants performed a visual search task with varying set sizes. Results suggested the difference between low and high literate groups in the functional visual field from the fovea to parafoveal areas indicating that reading-writing ability is not limited to verbal domain but can extend to nonverbal attentional processes (Olivers et al., 2014).
Literacy and cognition: fMRI studies
In this project, illiterate participants were trained to read and write Hindi (Devnagri) script for six months. Resting-state fMRI data were acquired for pre and post-training reading and writing the Devnagari script. This research suggested that training for reading and writing for six months could form new neural connections (Hervais-Adelman et al., 2019). This research also suggested that brain plasticity is possible in a mature brain even with six months of training to learn reading and writing. One of the significant findings of this project suggested that reading and writing skill does not negatively affect the visual processing regions related to nonorthographic material processing (Skeide et al., 2017).
[Reference: Olivers, C. N. L., Huettig, F., Singh, J.P. & Mishra, R.K. (2014). The influence of literacy on visual search. Visual Cognition, DOI: 10.1080/13506285.2013.875498
Hervais-Adelman, A., Kumar, U., Mishra, R. K., Tripathi, V. N., Guleria, A., Singh, J. P., Eisner, F., & Huettig, F. (2019). Learning to read recycles visual cortical networks without destruction. Science Advances, 5(9), eaax0262. https://doi.org/10.1126/sciadv.aax0262
Skeide, M. A., Kumar, U., Mishra, R. K., Tripathi, V. N., Guleria, A., Singh, J. P., Eisner, F., & Huettig, F. (2017). Learning to read alters Cortico-subcortical crosstalk in the visual system of illiterates. Science Advances, 3, e1602612. doi: 10.1126/sciadv.1602612]
We are interested to understand the link between literacy and cognition, more specifically the interaction between literacy and non-verbal cognitive processes.
Cognitive Development
Development of attention and control processes
In the past Gupta & Kar (2009) have examined normative development of attention and control processes such as task switching and error monitoring using the attentional disengagement task (see Figure 13) has shown transition in development of control processes like set shifting after 9 years of age until 11 years of age in terms of reduction in switch cost and error monitoring (see Figure 14).
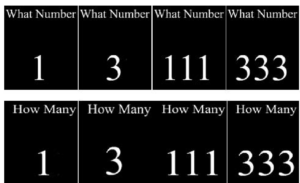
Figure 14. Presenting the sample stimuli of the switching task
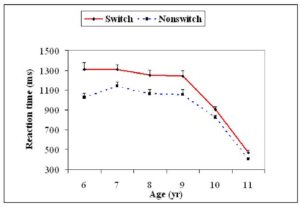
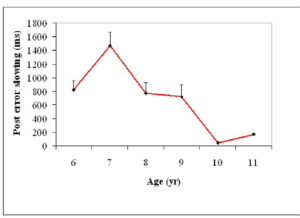
Figure 15. Performance on the switching task with respect to attentional shift (left) and post error slowing as a measure of error monitoring (right).
[Reference: Gupta, R. & Kar, B. R. (2009). Development of attentional processes in children with ADHD and normally developing children. Progress in Brain Research, 176, 259-276].Development of affective control
Our current studies examine affective conflict adaptation as a measure of proactive control across positive and negative emotions in children and adolescents as predicted by child’s temperament and family expressivity. We find that affect valence influences proactive control with negative emotions resulting in stronger adaptation effects, observed consistently among adolescents and happy affect enhances proactive control in children.
Development of cognitive-social-emotional competence
Our current projects on cognitive development investigate shifts in proactive-reactive control strategies from preschool to middle childhood and its relationship with academic and learning outcomes. The other ongoing project aims to investigate the development of social emotional competence and its relationship with variations in executive function profiles, temperament and learning/academic outcome in children using behavioural paradigms.
Cognitive and affective ageing
We also study the effect of cognitive aging on control processes and emotion regulation in middle aged and older adults. We found that the effects of aging in older adults may result in difficulties in early stages of perceptual processing, inhibitory control and unsuccessful suppression of irrelevant stimuli leading to difficulties in memory consolidation. Our recent studies on emotional ageing find a shift in affective bias towards positive affect in middle aged and older adults through implicit adjustments in cognitive control (Nigam & Kar, 2021) (see Fig 15). We have examined emotional aging across the life span using behavioural experimentation as well as experience sampling. Our current studies focus on understanding the mechanisms and implications of such positive affect bias for cognitive aging with behavioural and fMRI studies.
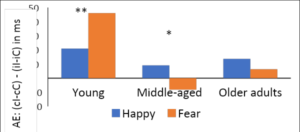
Figure 16. Shows that the magnitude of adaptation effect is more for positive affect among Middle aged and older adults compared to young adults. (AE: Adaptation effect).
[Reference: Nigam, R. & Kar B. R. (2021). Conflict monitoring and adaptation to affective stimuli as a function of ageing. https://doi.org/10.1007/s10339-021-01042-5]Cognitive Disorders
Our studies on dyslexia have looked at phonological awareness in Hindi and English language, central auditory processing deficits and script dependent vs central processing deficits in dyslexia. We have also been a part of a large-scale project on development and standardization of a multilingual tool for Dyslexia assessment for language of India (DALI) (Rao et al., 2021). One of our major projects on children with ADHD offers a classification system based on cognitive-motivational functions that are specifically impaired in ADHD children. This study indicates that response inhibition, error monitoring, task switching, and motivational style show considerable promise for the differential classification of ADHD (see Figure 16) (Gupta, Kar & Srinivasan, 2011).
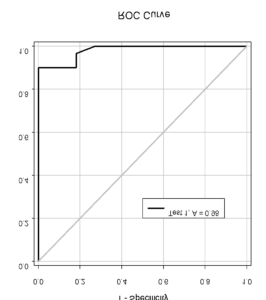
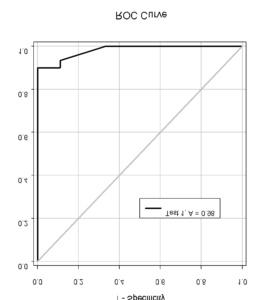
Figure 17. Short delay reward (left) and long delay reward (right) as a measure of delay aversion (motivational style deficit) is one of the best classifiers for children with ADHD compared to typically developing children and those with oppositional defiant disorder (9 years of age).
[Reference: Rao, C., Sumathi, T. A., Midha, R. Oberoi, G., Kar, B. R., Khan, M., Vaidya, K., Midya, V., Raman, N., Gajre, M., & Singh, N. C. (2021). Development and Standardization of the DALI-DAB (Dyslexia Assessment for Languages of India – Dyslexia Assessment Battery). Annals of Dyslexia. doi: 10.1007/s11881-021-00227-z]Gupta, Kar & Srinivasan, 2011 Gupta, R., Kar, B.R., & Srinivasan, N. (2011). Cognitive-motivational deficits in ADHD: Development of a classification system. Child Neuropsychology, 17, 67-81]
Action Planning, Control and Consequence
- Influence of spatial and temporal attention on executive control
An object at the attended location is perceived better, and a movement to the object is generated faster. It has also been reported earlier that once attention is deployed to an object, visual system takes a few hundreds of milliseconds to redeploy to another object. Detectability of a novel object appearing within this period is found impaired. Thus, a common consensus is that attention, in both spatial and temporal domain, plays key role in perception, target selection and planning a motor action to the target. However, if the context in which the movement has been planned alters, inhibition may be more essential for survival than execution of the movement. The fundamental questions remain, whether the same attention that facilitates visual perception also participates in executive control of motor action, or attention for perception and action are dissociable? How does the detectability of a perturbation in the visual world affect action control? Dr. Ray and his team seek answers to these questions using a set of novel variants of the oculomotor countermanding task in which the spatio-temporal allocation of attention is manipulated, and subjects are asked to inhibit impending rapid eye-movements, also called saccades, in response to an infrequent ‘stop’ signal.

Figure 18: Laboratory set-up for tracking gaze positions and the size of the pupils of the eyes.
A pool of young adult participants selected a target to look at by discriminating a go-cue in a stream of English letters, called rapid serial visual presentation (RSVP) task. Infrequently participants cancelled saccade and reported whether they saw the stop-signal in the stream. The go-cue and stop-signal both were embedded in a stream of irrelevant stimuli presented in rapid succession. Participants exhibited difficulty in detection of the stop-signal when presented almost immediately after the go-cue. Dr. Ray and his team found a robust relationship between the detectability of the stop-signal and the odds of saccade inhibition (see figure below, left). Saccade latency increased exponentially with the maximum time available for processing the stop-signal before gaze shifted (see figure below, right). A model in which the stop-signal onset spontaneously triggered deceleration in progressive saccade planning with magnitude proportional to its detectability accounted for the data.
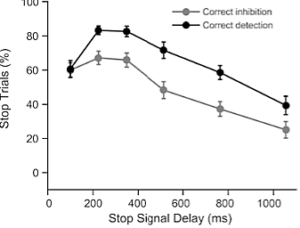
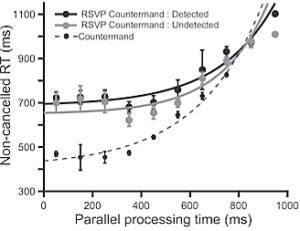
Figure 19: (Left) The ability to inhibit planned movement depended on the ability to detect the stop-signal. (Right) Exponential in reaction time with the duration of processing both go- and stop-signal simultaneously (i.e., parallel processing time).
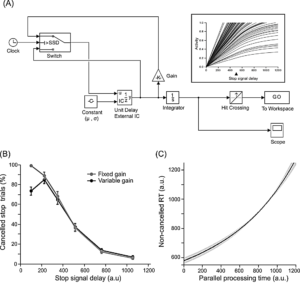
Figure 20. (A) Matlab Simulink model of countermanding; (B – C) Simulated data (compare with empirical data shown above)
[Reference: Indrajeet I, Ray S (2019). Detectability of stop-signal determines magnitude of deceleration in saccade planning. European Journal of Neuroscience, 49(2), 232-249.]- The rate of change in pupil size is an indicator of speed of movement planning.
The pupils of the eyes automatically constrict in light and dilate in dark to limit the amount of light entering the eyes. Many cognitive processes including decision making and movement planning can influence pupil size. The frontal eye field (FEF) in the cortex and subcortical superior colliculus (SC) critically participate in planning eye-movement and send projections to the parasympathetic and sympathetic divisions of the autonomic pupillary light response (PLR) network, respectively. While the former network causes constriction of pupils the latter is responsible for dilation.
A recent study in Dr. Ray’s lab has demonstrated a tight coupling between the oculomotor system that plans eye movements, and the autonomic system that controls pupillary aperture size. This finding suggests that the brain is capable to adjust pupil size during preparation for eye movements to proactively counter transient changes in the intensity of light entering the eyes. They endorse that the pupil size is a potential biomarker for action planning.
Figure 21. Simulation of a neuro-mechanical model of PLR accounted for a robust inverse relationship between the reaction time of eye movement and the rate of pupil constriction.
[Reference: Pandey P, Ray S† (2021) Pupil dynamics: A potential proxy of neural preparation for goal-directed eye movement. European Journal of Neuroscience, 54(7), 6587 – 6607]- Inhibitory motor control in Tourette disorder patients
Tourette disorder (TD), represents a relevant model of inhibition impairment due to its major clinical sign – uncontrolled tics. Previous studies that attempted to contrast inhibitory control of impending motor action in Tourette disorder and healthy humans using traditional ‘race’ model reported mixed results. Race model assumes that two independent processes, called GO and STOP, compete with each other while rising to reach a fixed threshold.
Dr. Ray and his collaborators used a pair of parameters derived from the ‘Cancellable Rise to Threshold’ or CRTT model (Indrajeet & Ray, 2019, 2020; Salinas & Stanford, 2013) to estimate inhibitory control in patients and healthy humans. A population of 53 TD patients and another population of 30 age-matched healthy humans participated in a countermanding task (Figure 21), where they had to cancel a movement in response to a stop-signal that appeared suddenly in some trials. Occasionally they failed to inhibit planned movement. Reaction times in those non-cancelled trials were plotted as an exponential function of the duration of simultaneous processing of go- and stop-signal. The rate of rise in the slope of the function in logarithmic scale estimated reactive control, while the intercept of the function on the ordinate estimated proactive control. Those novel metrics were referred to as ‘log-attenuation rate’ and ‘proactive delay’.
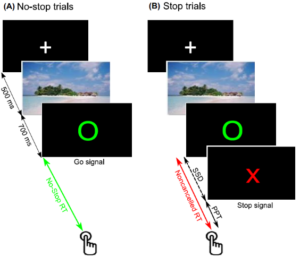
Figure 22: Schematic of stop-signal task
Neither the stopping behavior nor the metric derived from race model was able to distinguish between TD and HC. On the contrary, our new method based on CRTT model distinguished contributions from proactive and reactive control in stopping a manual movement. While the ability to postpone an action by TD was comparable to HC, the log-attenuation rate that estimates the strength of deceleration in GO process following the stop signal was significantly less in TD than HC (Figure 22).
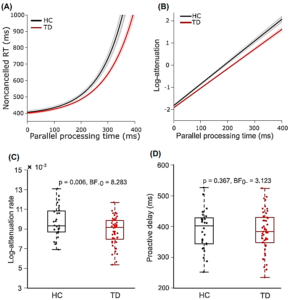
This study challenged the conventional method of estimation of inhibitory control in healthy humans and patients. The primary advantage of this method is that it can tease apart reactive and proactive control. This study has resolved a long-standing conundrum: why the impairment in the inhibitory control in TD patients could not be identified despite their atypical neural networks critical for executive control? In summary, this study showed that the reactive control was weak in TD patients in comparison to healthy age-matched adults, which they rectified by intact proactive control.
[Reference: Indrajeet, I., Atkinson-Clement, C., Worbe, Y., Pouget, P., & Ray, S. (2022). Compromised reactive but intact proactive inhibitory motor control in Tourette disorder. Scientific reports, 12(1), 1-11.]Future direction
Electroencephalography (EEG) on humans: Previous studies have shown diminished or absent N2pc component of ERP, when a target goes unnoticed in a rapid visual presentation, but existence of N400 component indicating its semantic representation. Comparison of performance, response times, and attention related modulation of EEG signals in different task conditions will help us to understand the dynamics of target selection and action planning.
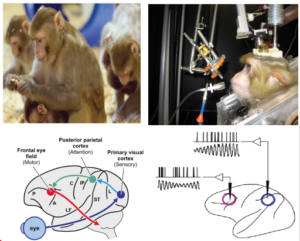
In vivo electrophysiology on non-human primates: Rhesus monkeys will be trained to perform cognitively demanding tasks similar to tasks described above. Simultaneously recorded spiking activity and local field potentials of neurons in the frontal eye field (FEF) and lateral intra-parietal (LIP) area of monkeys will be examined for synchrony and causality. Activity of recorded neurons and the simulated Bayesian neurons will be compared to understand the computations in this network. The outcome of this project will improve our understanding of the functional role of parietal and frontal cortices in eye-hand coordination and decision-making.

Learning and Memory
In the past some studies were conducted to understand skill acquisition. The behavioural and neural findings (functional MRI) suggest sequence chunking and its underlying mechanisms. The fMRI findings suggest that Fronto-parietal network brain activations are due to increased long-range sequence complexity, which results in long-range sequence chunking (Pammi et al., NeuroImage, 2012). Miyapuram, Singh, Bapi & Pammi (2011) proposed a cognitive model that learns visuo-motor sequences using a limited capacity working memory (WM) would need to optimize in two different ways depending on the amount of information to be processed at any instance of time.